|
†
WYBIERZ RÓD:
.
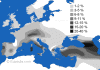
PIERWOTNIE NIEINDOEUROPEJSKIE
HAPLOGRUPY Y-DNA w EUROPIE,
zwłaszcza staroeuropejska I (I1 i I2)
(mitochondrialne mtDNA - zob.
na dole)
Kiedy
indoeuropejskie populacje R1a i R1b zaczęły w polodowcowym
mezolicie, neolicie lub epoce brązu zasiedlać Europę, zastały tu
zapewne bardzo
rzadkie osadnictwo ludności starszej, przed-indoeuropejskiej,
która przeżyła okres wielkiego zlodowacenia. Pod względem Y-DNA
to haplogrupa I (I1 i I2).
Tak więc pod
względem pochodzenia ojcowskiego ludność Europy nie jest
zupełnie jednolita. Populacje pierwotnie indoeuropejskich haplogrup:
"słowiańskich" R1a1a1 i "italo-celtyckich" R1b1a2, czyli twórców języka
praindoeuropejskiego,
stanowią około 70 procent Europejczyków. 20 procent to
chyba jedyna przedindoeuropejska haplogrupa I (I1 i I2),
czyli prawdopodobnie jacyś "Wenetowie".
Pozostałe 10 procent to zwłaszcza bliskowschodnie J1 i J2, północnoafrykańsko-egejska
E, północnosyberyjska i ugrofińska N, małoazjatycka/kaukaska G i
hinduska/cygańska H.
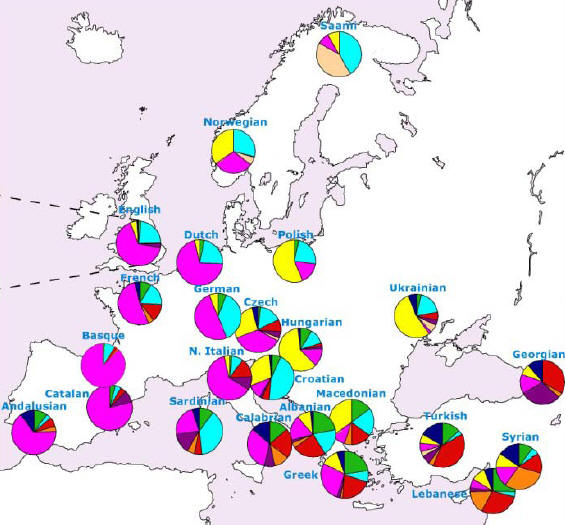 |
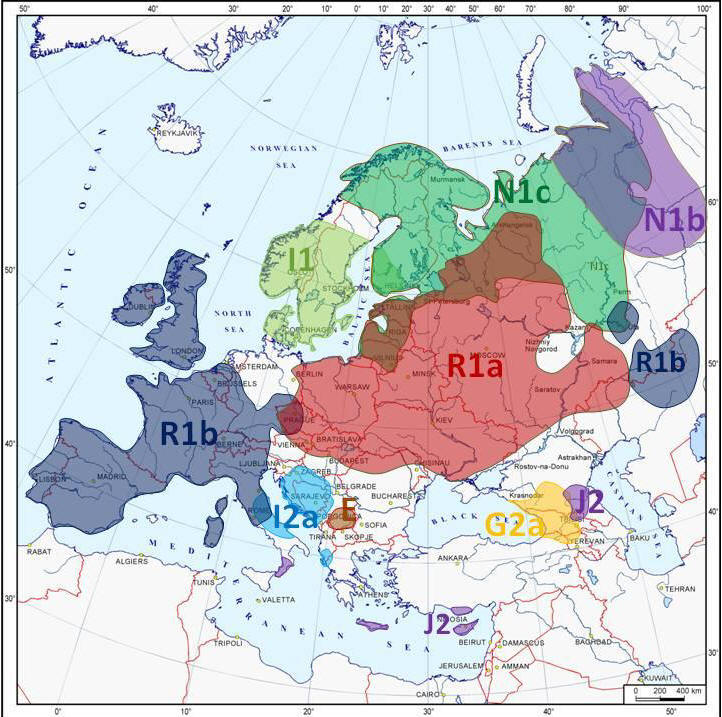
Podstawowe genetyczne rody YDNA w
Europie
Europa Środkowo-Wschodnia to głównie
R1a1a1.
Europa Zachodnia to głównie
R1b1a2.
Europa Północno-Wschodnia to głównie
N1c.
Góry Dynarskie i Sardynia to I2a.
Kraje nadbałtyckie to nadto I1.
Europa Południowa
to nadto G, J2 i K.
dna-forums |
http://www.nottingham.ac.uk/-sczsteve/Gothenburg_13Oct2011.pdf
HAPLOGRUPA I
(tj. I1 i I2)
(to prawdopodobnie przedindoeuropejscy Enetowie/Wenetowie/Wenedowie?).
Ważne publikacje:
A.Klosow, Gaplogruppa I, Wiestnik
Rossijskoj Akademii DNK-genealogii, 3/1 2010 (s. 96-158)
http--www.lulu.com-items-volume_67-8049000-8049755-2-print-8049755.pdf,
str. 96-158.
http://www.familytreedna.com/public/I2aHapGroup/default.aspx?section=yresults
(mapka części populacji haplogrupy I1 i I2 w FalilyTreeDNA)
Wiestnik, maj 2011(ros.):
http://www.lulu.com/items/volume_70/10588000/10588664/1/print/10588664.pdf,
str. 968-1114
Forum Rodstwo (ros):
http://www.rodstvo.ru/forum/index.php?showtopic=1814&st=20
Ken Nordetvedt:
http://knordtvedt.home.bresnan.net/Tree%20and%20Map%20for%20Hg%20I.pdf
:
tam komentarz (s.1), mapa migracji (s.2), drzewo i datowanie
(s.3). Autor jest wybitnym znawca haplogrup I1 i I2.
Zob. też: Wikipedia Haploroup I; Haplogrupa I oraz Eupedia
...
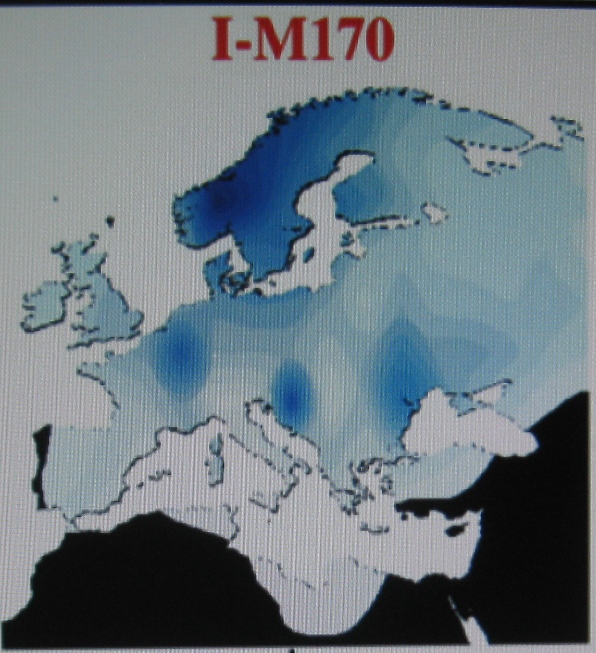
Między 48.000-40.000 lat temu pojawiła się w Europie
pierwsza populacja z gatunku homo sapiens-sapiens ("człowiek
współczesny"), zidentyfikowana po raz pierwszy w Cro-Magnon
w poludniowo-zachodniej Francji, zapewne nosiciel zespolonej
haplogrupy IJ. Był to tzw. "człowiek kromanioński" (Zachodznia
Francja), albo "człowiek bohunicki",
zidentyfikowany w miejscowości Bohunice k.Brna (Czechy) oraz
w kilku miejscowościach Europy południowej (Włochy),
środkowej (Bułgaria, Morawy i Polska-Dzierżysław) lub
wschodniej, lub jako "człowiek z Kostienek" nad Donem z
około 30000 lat temu. Nie znaleziono dotąd ich
wyrobów-narzędzi, ale np. przez porównanie z człowiekiem z Ahmarian (Turcja) uznano bohunicjan za ludzi współczesnych,
ich kulturę materialną określono jako proto-oryniacką, a
jako ojczyznę ich pochodzenia uznano Bliski Wschód. Z powodu
wystąpienie około 40.000 lat temu zjawiska "zdarzenie
Heinricha4", spowodowanego chyba wulkanicznym Ignimbrite,
którego skutkiem było gwałtowne, trwające kilkaset lat
zanieczyszczenie i oziębienie atmosfery, człowiek bohunicki
mógł prawie całkowicie wyginąć.
Prawdopodobnie po tym zdarzeniu, około
38.000 lat temu pojawia się już człowiek "klasycznej"
kultury oryniackiej. Kultura ta znana jest w całej Europie
35.000-29.000 lat temu, także w Polsce, np. w jaskini w Obłazowej koło Nowego Targu (zob.
Dyskusja n.13).
W sumie to około 20% dzisiejszych mieszkańców Europy.
Pod względem genetycznym
człowiek oryniacki odznaczał się najprawdopodobniej
zespoloną haplogrupą IJ-M429,
powstałą zapewne w Mezopotamii, około 50000 lat temu. Stamtąd, według
przypuszczeń A.Klosowa, poprzez Kaukaz dostał się na ziemie nad Wołgą i Donem. Jego śladem może być
tzw. człowiek z Kostienek (z około 30.000 lat). Ale nie istnieje ta mutacja do dziś, gdyż jej populacja mogła wyginąć w czasie ostatniego wielkiego
zlodowacenia (MGL, od około 20.000 lat temu).
Według I.
Rożanskiego (Rodstvo.ru, maj 2011) w części europejskiej populacji IJ
około 28.000 lat, może w rejonie Bałkanów, a według Ken
Nordvedta w pn.-zach. Azji Mniejszej, rejon Bosforu, około 24000
lat temu, wyłoniła się haplogrupa I-M170. Zapewne trudne warunki
egzystencji sprawiły, że tylko nieliczni przedstawiciele tej
haplogrupy zdołali się przecisnąć przez demograficzne "szyjki
butelki" w okresie ostatniego wielkiego zlodowacenia i mezolitu,
dlatego przodkowie (MRCA) głównych ich dzisiejszych populacji
datowani są przeważnie na czasy neolitu i późniejsze. Natomiast około 23.000 lat
temu z tej samej zespolonej haplogrupy IJ wyłoniła się
haplogrupa J-M304. Ta zapewne przed wielkim zlodowaceniem uszła ona na Kaukaz
i prawdopodobnie tam wydała dwie swoje podgrupy:
około 20.000 lat temu J2-M172 i około 16.000 lat temu J1-M267.
Były to
populacje staroeuropejskie, przedindoeuropejskie, językowo
spokrewnione zapewne z językiem makrogrupy afroazyjskiej z
Bliskiego Wschodu. Zapewne one zapoczątkowały i rozwijały od około
28.000 do 22.000 kulturę grawecką (znaną także z jaskini Obłazowej
na Podtatrzu),
oraz młodsze jej pochodne, np. polodowcową kulturę
magdaleńską, spotykaną także w kilku regionach Polski
Południowej. Prawdopodobnie ta haplogrupa I, rozproszona po Europie
w kilku genetycznych podgrupach, tworzyła populacje, zwane
w starożytności i średniowieczu Enetowie/Wenetowie/
Wenedowie i podobnie (zob.
Dyskusja n.3).
 |
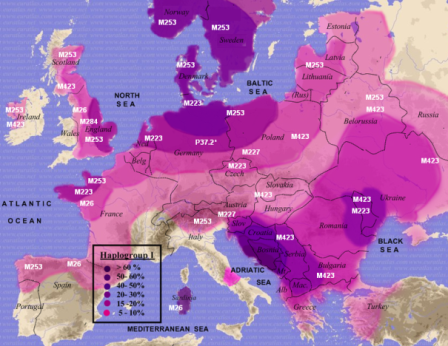
Przedindoeuropejska haplogrupa I
i jej podgrupy w
Europie, wg.
Chiaroni et al. 2009
1. Skandynawia, Niemcy, Nadrenia:
to "germańskie" hg. I1 (M253) i
I2b1 (M223).
2. Sardynia, Płw. Iberyjski, Irlandia: to hg I2a1
(M26).
3. Chorwacja i Ukraina:
to dynarska, "słowiańska" hg I2a2 (M423). |
9-1A) Haplogrupa I1, głównie
w ludach germańskich.
Z ojcowskiej haplogrupy I
może gdzieś na Bałkanach około 22000 lat temu, a więc w okresie jeszcze przed LGM, wyłoniła
się w drodze mutacji M253 haplogrupa I1
("i-jeden"); jej populacja, przetrwawszy zlodowacenie,
rozprzestrzeniła się z terenu północnych Niemiec na Jutlandię,
Skandynawię, Wyspy Brytyjskie i ziemie podalpejskie. Zasadniczy czas jej rozwoju i ekspansji przypada
jednak dopiero na drugie tysiąclecie p.n.e. Dziś w Szwecji stanowi
około 42% ludności, w Norwegii 36%, Islandii 33%, Danii 30%,
Finlandii 28%, a w Niemczech ponad 20% (zob. w Eupedii).
Udało się ustalić, że wspólny przodek haplogrupy I1 w Anglii, Irlandii i Szkocji żył
około 1425 przed Chr; w Skandynawii - 1375; w Europie
kontynentalnej od Hiszpanii do Słowacji - 1425; we Europie
Wschodniej łącznie z Polską - 1225; tak samo w Niemczech; Bliski
Wschód - 1475 przed Chr. Ekspansja tej haplogrupy czasowo
kojarzy się więc z ekspansją słowiańskiej haplogrupy
R1a1a.
Tu
dygresja o powstaniu Germanów.
Choć haplogrupa I1 pojawiła się w
Skandynawii wcześniej, tworząc tam wielkie starożytne kultury
przedneolityczne, to jednak około 1300-1400 lat p.n.e. na Półwyspie
Skandynawskim przebywała ich bardzo mała wspólnota. Jeden bowiem mężczyzna tej haplogrupy
(MRCA)
około 1375 p.n.e. dał tam początek
wszystkim późniejszym populacjom nordyckim tej haplogrupy.
Nieco później pojawiło się tam także i plemię "słowiańskie" z hg R1a1a1, oraz północno-zachodni oddział
z mutacją M223, a w ramach
migracji z celtyckiej kultury Hallstatt pod Alpami ok. 700 p.n.e. dotarły tam
plemiona haplogrupy R1b1a2. I właśnie z tych trzech grup: I1+I2a2, R1b1a2 i
R1a1a1 na styku Skandynawii i północnych Niemiec powstali Germanowie. Dziś wśród
nich około
40% to haplogrupa I1/I2, około 40% - R1b1a2 i około 20% - R1a1a. Tak więc rasistowskie myślenie
niektórych dawnych Niemców o czystości rasowej Germanów zostało przez dzisiejszą
genealogię genetyczną zupełnie skompromitowane.

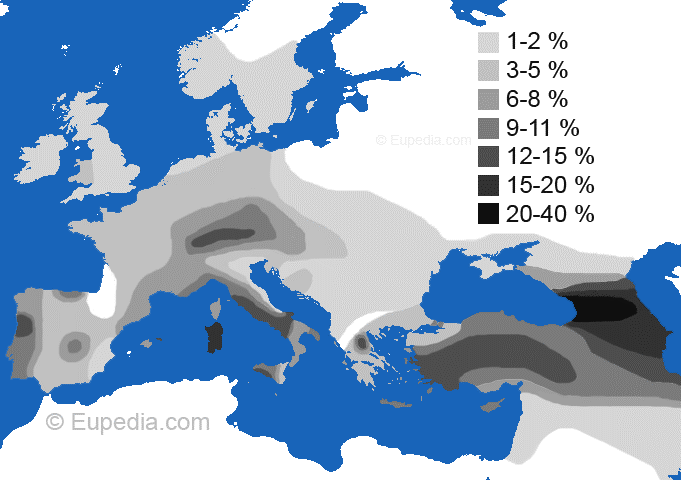
Populacja hg I i I2;
główne podgrupy |
Na lewo:
Przedindoeuropejskie haplogrupy I i I2 i ich główne podgrupy.
Na prawo:
Podgrupa
bałkańskosłowiańska
I2a1-M423,
w dzisiejszym jej zagęszczeniu:
Bałkany Zachodnie (Bośnia-Hercegowina)
- miejsce genezy - do 75% (mapy z Wikipedii) |
 |
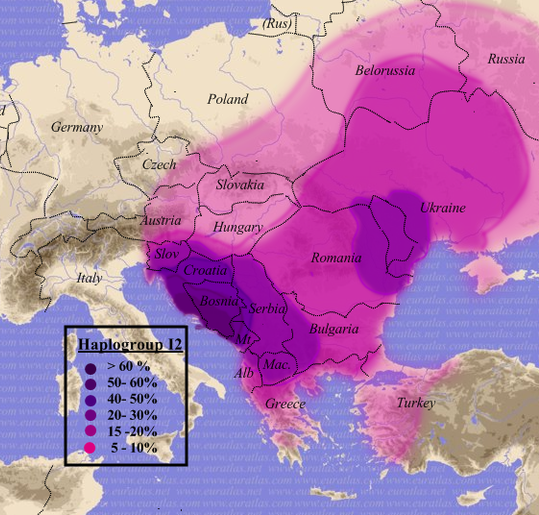
9-1B) Haplogrupy I2, I2a1 -
dziś głównie
słowiańskie.
W populacji hg
I po mutacji M438, około 25.000-20.000 lat temu, a więc także przed
ostatnim zlodowaceniem, wyłoniła się haplogrupa I2-L460,
a w niej później podgrupa I2a1a-M26,
zasiedlające głównie Sardynię, oraz podgrupa
I2a1b-M423,
która
w swoim czasie, wraz ze Słowianami
(R1a1a1), zdominowała niektóre rejony bałkańskie i
środkowo-wschodniej Europy: Chorwacja (i jej niektóre wyspy) 42%, Bułgaria 33%, Serbia 27%, Rumunia
17%, Węgry15%, Białoruś 25%, Ukraina
12%. pd.-zach. Rosja 11%, Polska 10%, Słowacja 10%); zob. w Eupedii. Nic dziwnego, że w
starożytności niektórzy autorzy nie umieli rozróżnić między
rdzennymi Słowianami (R1a1a1) a zeslawizowanymi "Wenetami" (I2a1b); już wtedy obie populacje
tworzyły zapewne jakby jeden etnos. To zapewne ci Wenetowie
mieli udział w tworzeniu
najstarszych kultur archeologicznych na stepowych terenach Ukrainy
i południowej Rosji, np. udział w Cucuteni-Tripolje, k. kurhanowej czy ceramiki sznurowej, zanim
nie dołączyła do niej młodsza, przybyła z zachodnich Bałkanów, znad Dunaju
lub od Karpat, populacja słowiańska R1a1a1.
Udało się ustalić pierwszych
wspólnych przodków dla poszczególnych populacji haplogrupy I2.
Okazała się ona, wg. Klosowa, bardziej niż h.g. I1 odporna na przeciwności
życia w tamtym czasie. I tak według niego: wspólny przodek
wszystkich populacji żujących dziś I2 żył 13.000 p.n.e. (przybył
tam zapewne później, po MGL); w Irlandii 14.200
p.n.e.; w Szkocji 14.800 p.n.e.; w Skandynawii 13000 p.n.e.;
Europa kontynentalna od Hiszpanii do Słowacji 1425 p.n.e.;
Niemcy 9675 p.n.e.; Europa Wschodnia łącznie z Polską 14200
p.n.e..
Tu dygresja o powstaniu Rusi.
W
ruskich Latopisach napisano: "Przyszli Waregowie. Nazywali
się Ruś... Mieli język ruski i słowiański". W XVIII w.
dopisano z fantazji, że Waregowie przyszli zza morza i byli
Normanami. Faktycznie to chodziło raczej o plemiona Rugów/Rusów,
żyjących wśród Słowian co najmniej od II w. w pasie miedzy
Bałtykiem a Morzem Czarnym i zmagających sie z naporem
germańskich Gotów. Ich genetyczne haplolgrupy to I1 (populacja
bałtycko-skandynawska) oraz I2a2 (populacja
południowosłowiańska). Ogólnie wśród dzisiejszych "Ruskich" hg
I2a1 to 15%, a I1 - 6,5% ludności, żyjących obok obok R1a1a -
48% i N1c - 14%. Zapewne z hg I1 był Ruryk, Olga, Włodzimierz
i inni protoplaści Rusi kijowskiej i nowogrodzkiej.
Zob. o tym: Klosow, Haplotypy południowych i bałtyjskich
ruskich Słowian: cztery plemiona? w:
Proceedings... t. II/5 (2009), s. 801-815 (por. A. Klosow).
Dygresja o powstaniu Bałtów.
Uważa
się, że etnos i język Bałtów (czyli zwłaszcza Prusów, Litwinów i
Łotyszy) wydzieliły się ze słowiańskich, o czym świadczą
niektóre wspólne innowacje językowe bałto-słowiańskie, i
to najpóźniej około 1000 lat przed Chr. Dziś w rejonie Prabałtów
spotykamy słowiańską haplogrupę R1a1a około 40% i
przed-indoeuropejskie: bałtycko-skandynawską I1 i bałkańską I2a2,
razem około 6% (Białoruś 25% I2a2). Można więc uważać, że
haplogrupy I1 i I2 (Wenetowie bałtyccy) uczestniczyły w tym
regionie w ewolucji słowiańskiego języka i wydzieleniu się jego
formy bałtyckiej (por. A.Klosov).
Haplogrupie I2a1 (M423), obok hg E1b1b1a2 (V-13), genetycy przypisują upowszechnianie
rolnictwa, przynajmniej w południowo-wschodniej Europie,
przejętego w okolicach wschodniego wybrzeża Adriatyku od przybyłej z Anatolii haplogrupy J2b2 (M241). Zob. Dyskusja
n.11.
Natomiast jej
siostrzana podgrupa I2a2 (M26),
wyłoniona około 19.000 lat temu,
zdominowała Sardynię (39%) oraz nielicznie przetrwała na
północnym zachodzie Europy (Kraj Basków, Bretonia, Anglia,
Irlandia). Populacjom haplogrup I1, I2a2 i R1a1a można by przypisać
dzieła kultury megalitów (kamiennych kręgów), w tym
słynnego Stonehenge w Anglii z około 2800 lat przed Chr.
UWAGA!
Obok Wenetów/Wenedów, stanowiących od starożytności wspólne
etnosy z Germanami lub Słowianami, naukowa literatura wymienia
także Wenetów iliryjskich na ziemiach północnej Italii (IX w.
przed Chr.), później uległych romanizacji, oraz Wenetów
Celtyckich, których etnonimy zostały zanotowane w Alpach
Szwajcarskich, w Zachodniej Francji i w Walii. Dopatrzyć się ich
można także pod nazwą "Enetoi" u Homera jako
uczestników wojny trojańskiej w Azji Mniejszej przed 1200
r. przed Chr. i osadników nad Dolnym Padem we Włoszech, a później - w Eneidzie Wergiliusza
(por. G. Jagodziński:
Zagadkowy lud - Wenetowie;
por. w
Wikipedia)
|
Populacje
I1 i I2 to prawdopodobnie tzw. Paleoeuropejczycy
(przedlodowcowi)
Haplogrupa I
wydzieliła się z ojcowskiej IJ może około 28.000
lat temu w Europie, na Bałkanach(?). Zapewne te populacje
były twórcami przedlodowcowej kultury graweckiej i polodowcowej
- magdaleńskiej. Przetrwały epokę wielkiego zlodowacenia
zapewne w refugium bałkańskim (np. hg
I2a1), lub kaukaskim (J, J2).
Już
w mezolicie i neolicie, I1 i I1, rozproszone były po całej Europie,
ale prawdopodobnie łączyła ich nazwa Wenetowie/Wenedowie.
Z napływowymi ludami,
nieco wyżej kulturowo zaawansowanymi populacjami R1a1a, a później także R1b1b2, tworzyli nowe,
krystalizujące się właśnie etnosy europejskie i niekiedy wpływały na kształt ich języków
indoeuropejskich populacji R1a (satemowych) i R1b (kentumowych); najwyraźniej swój udział zaznaczyli w
powstaniu etnosu germańskiego, dając mu zarazem znaczną część
swojego zasobu językowego (stąd określanie języka germańskiego
mianem kreolski). |
O obecności
haplogrup I1 i I2 w Polsce i w rejonie Dunajca,
zob.
Pozostałe mniejsze haplogrupy, pierwotnie
nieindoeuropejskie, w Europie
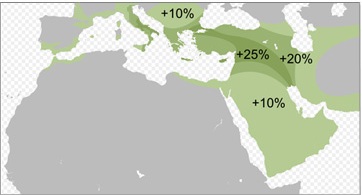
HAPLOGRUPA J2a i J2b, pierwotnie bliskowschodnie.
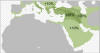
Haplogrupa
J2 jest dość powszechna w Europie południowej
i południowo-wschodniej, zwłaszcza w rejonie Grecji
(Turcja 33%, Kreta 32%, Albania 28%, Grecja 27%, Rumunia 24%,
Italia 20%). Powstała ona zapewne w rejonie południowego Kaukazu
i wnet upowszechniła się w rejonie tzw. Żyznego Półksiężyca (uprawne
tereny od
Zatoki Perskiej, przez Kurdystan, Syrię, Liban i Palestynę do
Delty Nilu).
W rejonie dawnej Fenicji
i Syrii haplogrupa J2a tworzyła wczesnoneolityczną kulturę impresso,
zwaną też Cardial Pottery (zob.
wyżej, mapa). Głównie więc populacji J2a Europa
zawdzięcza przeniesienie początków ceramiki na początku
neolitu (a więc przed 5500 lat przed Chr.) wzdłuż południowych
wybrzeży Europy, aż do Hiszpanii. Zaś do przejęcia i upowszechniania
rolniczego neolitu (m.in. w
kulturze LBK) po Europie przyczyniły na pewno populacje obydwu
europejskich haplogrup
R1a i R1b.
Natomiast
bratnia haplogrupa J1 zdominowała głównie kraje arabskie i
semnickie, promieniując także i do Europy.
O obecności
hapologrup J1 i J2 na polskiej ziemi i w rejonie Dunajca,
zob.
(W moim rodzie haplogrupą J2a4ha*
wyróżniał się
Wojciech Szlaga z Łukowicy)
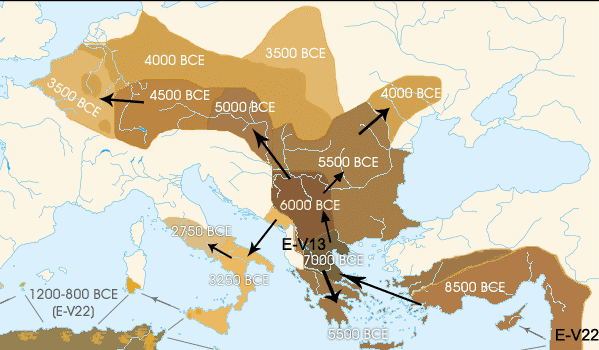
HAPLOGRUPA E1b1b1a2 (V-13)
(południowoeuropejska).
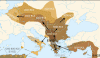
Północno-wschodnia afrykańska
populacja haplogrupy E, bardzo stara, bo powstała
ok. 52.000 lat temu i żyjąca głównie w północno-wschodniej i północnej Afryce
(zwł. Egipt i Etiopia), przybyła głównie przez małoazjatycką Anatolię
do Grecji (jako Fenicjanie?), w okresie przed-rolniczym,
około 7000 lat przed Chr. Występuje
zwłaszcza na południowych wybrzeżach Europy jako haplogrupa E1b1b1a
(M78). Z niej, już na terenie Europy, wyłoniła się rdzennie
europejska haplogrupa E1b1b1a2 (V-13).
Populacje E znajdują się: Grecja 27%, Serbia 24%, Albania 22%,
Portugalia 17%, Bułgaria 12%, Włochy 11%, Słowacja 11%.
Haplogrupie E1b1b1a2 (V-13)
nieśmiało przypisuje się przyniesienie do Europy elementów kultury znad
Nilu oraz - obok hg I2a2 (M423)
- upowszechnienie na Wybrzeżach Śródziemnomorskich rolnictwa,
przejętego od przybyłej z Anatolii w Azji Mniejszej haplogrupy J2b2 (M241).
O obecności haplogrupy E-V13 w
Polsce i w rejonie Dunajca,
zob, oraz wśród
Karpato-Rusinów (Łemków).
HAPLOGRUPA F
|
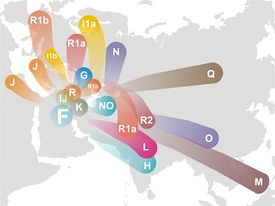
Ojczyzna i ród haplogrupy F według FtDNA |
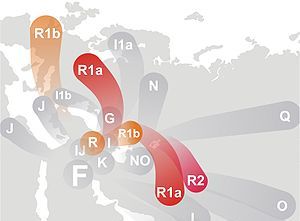
Ojczyzna i ród haplogrupy F według Wikipedii |
Haplogrupa
F, do niedawna w Europie niezauważana, bo bardzo rzadko
występująca, została zidentyfikowana przez
W. Haak et al. 2010 w dwóch z trzech szkieletów kultury
ceramiki wstęgowej z cmentarza w Derenburg (Saksonia, Niemcy).
Szkielety datowano na lata 5247 i około 5000. Kultura ceramiki
wstęgowej jest pierwszą europejską szeroką kulturą ceramiki i z
nią wiążą się początki rolnictwa w Europie, przeniesionego z
Bliskiego Wschodu. Można jednak przyjąć przypuszczać, że
haplogrupa F, która przecież powstała około 48000 lat temu,
miała dość czasu, by zabłądzić do Polski daleko przed neolitem i
przyjąć rolnictwo od przybyszów G2a3 lub R1a1a jako autochton
Derenburga.
HAPLOGRUPA G2a, pierwotnie kaukaska.
Video o dolmenach na Kaukazie wśród ludności haplogrupy
G2a
http://www.youtube.com/watch?feature=player_embedded&v=KrIrdIyHBNU
Sporadycznie i w niewielkiej liczbie występuje w Europie ludność
haplogrupy G2a. Przybyła ona z Azji
Mniejszej, z rejonu Kaukazu, głównie może z populacji Ormian lub
Gruzji (tam około 30-40%) i
sąsiednich populacji oraz Turcji (kilkanaście procent). W
znacznym procencie zaludniają dziś Sardynię (15%). Poniżej 10%
zaludniają Szwajcarię, Austrię, południowe Niemcy i Czechy.
Haplogrupę G2a3 zidentyfikowano
obok F* wśród szkieletów z cmentarza kultury ceramiki wstęgowej
w Derenburg w Niemczech.
HAPLOGRYPA N1c i N1b, ugrofińskie.
Północnosyberyjskie
i uralsko-fińskie populacje to
haplogrupa N
(na drzewie genealogicznym hg N to jakby daleka kuzynka haplogrupy R).
Haplogrupa N wyłoniła się we wschodnich Chinach około 10.000 lat
przed Chr., a jej podgrupy przez rejon Ałtaju dotarły do ziem Syberii
podbiegunowej, a stąd około 4-5 tysięcy lat przed Chr (AK) do
północnowschodniej Europy.
Licznie reprezentowana
podgrupa N1c (M46/Tat)
datowana jest na około 1500 lat przed Chr., głównie w północnowschodnich rejonach Europy
od Uralu i górnej Wołgi po północną Skandynawię (Suomi-Lapończycy).
I tak: Finlandia 58%, Litwa 42%,
Łotwa 38%, Estonia 34%, Rosja 23%.
Wśród Węgrów, których język ma
pochodzenie ugrofińskie, tej haplogrupy jest tylko 1% (co ujawnia, że w
zdobyciu ziem słowiańskich nad Dunajem w końcu IX w.
uczestniczył tylko niewielki oddział rdzennych Madziarów, albo
wytępiły ich średniowieczne najazdy mongolskie i tureckie). Daleko mniej
liczna występuje wśród nich także siostrzana
haplogrupa N1b (M43).
HAPLOGRUPA H1a, hindusko-romska
Haplogrupa H1a to głównie pochodząca z Indii (Pendżab) populacja
europejskich Cyganów i pokrewnych im plemion (np. Romów). Ich
średniowieczne przybycie do Europy datowane jest na około IX wiek. Ich język
należy jednak do grupy indoirańskiej z półwyspu Indyjskiego.
Powyższe haplogrupy, choć genetycznie nieindoeuropejskie, w znacznej
mierze przyczyniły się nie tylko do powstania dzisiejszego
kształtu języków indoeuropejskich w Europie, ale także całego
bogactwa kultury duchowej i materialnej Europy, każda z nich w
swoim zakresie i według własnych możliwości populacyjnych i
czasowych.
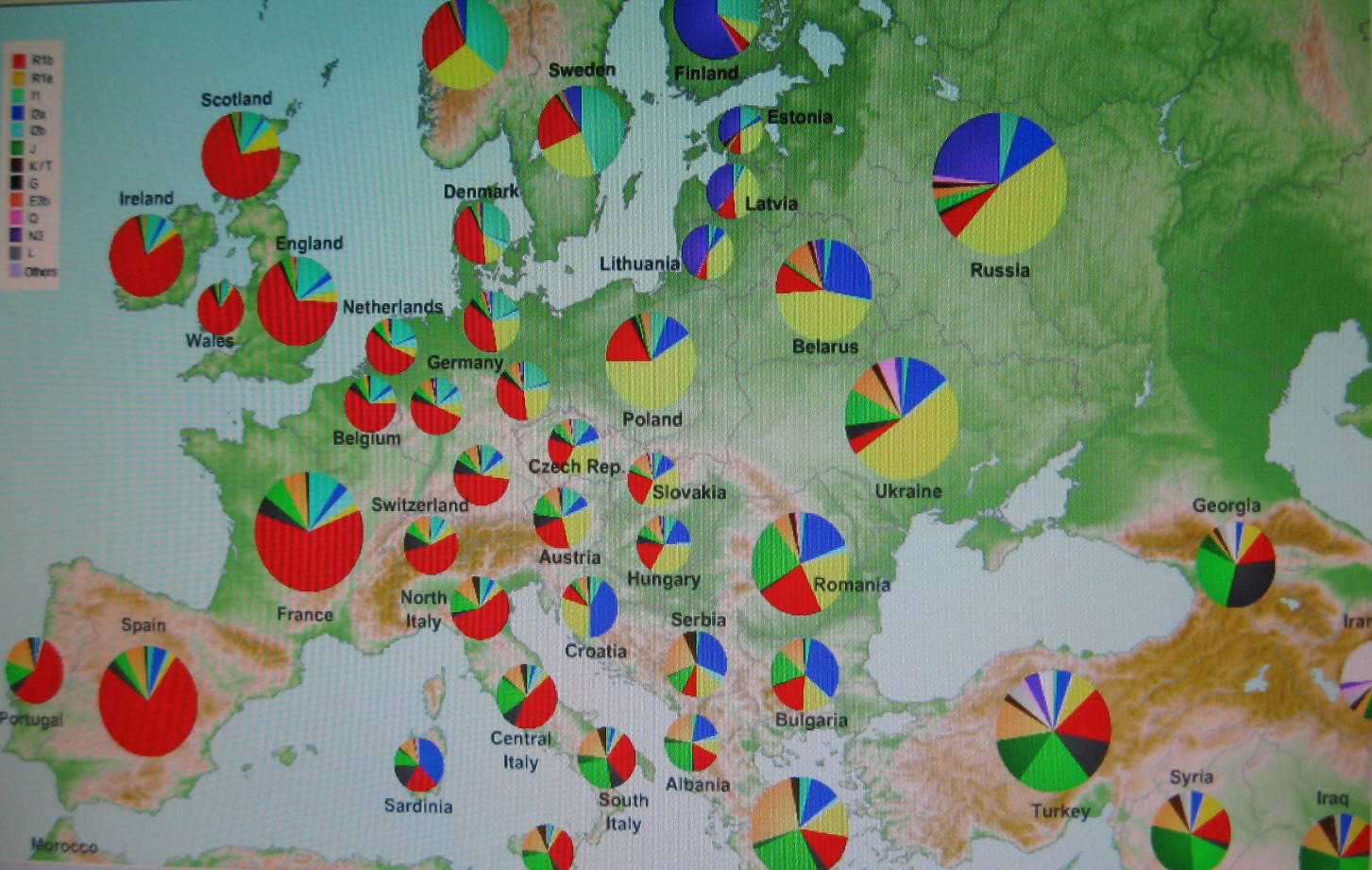
Główne rody Y-DNA w Europie
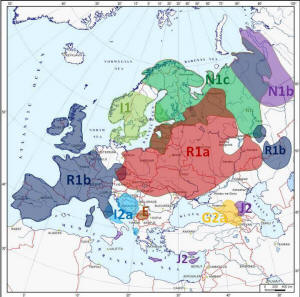
Szczegółowe
rozmieszczenie rodów Y-DNA w Europie
(według portalu
Eupedia.tabl)
|
 |
Kolor
żółty
R1a1a
(wschodnioeuropejski, słowiański);
czerwony R1b1b12
(zachodnioeuropejski, celtycko-germański);
modry
I1
(nordycko-germański);
granatowy I2a
(bałkańsko-słowiański);
błękitny I2b
(bałkańsko-niemiecki), pomarańczowy
E
(północno-wschodnioafrykański i grecki);
fioletowy N
(uralsko-fiński, północnorosyjski), zielony
J1
(semicki) i
J2
(bliskowschodni), czarny
G
(kaukaski). |
MITOCHONDRIALNE DNA
WSKAZUJe MATCZYNE POCHODZENIE CZŁOWIEKA
DNA
mitochondrialne, czyli mtDNA, to materiał genetyczny znajdujący się w
rejonie pozajądrowym komórek. Białka kodowane przez mtDNA to część
białek łańcucha oddechowego (regulują
energetykę komórki i całego organizmu;
odpowiadają też za ciepłotę ciała).
DNA
mitochondrialne są wykorzystywane w genealogii genetycznej i
medycynie sądowej. Służą do tego niekodujące fragmenty (regiony)
genomu mitochondrialnego o wysokim zróżnicowaniu i zmienności u poszczególnych ludzi.
Szybkozmienny region HVR1
obejmuje sekwencję 16001-16568, a region HVR2 - sekwencję 001-574.
Porównanie DNA mitochondriów
pochodzących od ludzi, wywodzących się z różnych grup etnicznych,
pozwoliło na obliczenie, kiedy żyła tzw. Ewa mitochondrialna -
kobieta, od której wywodzą się wszyscy współcześni ludzie, a właściwie ich genom mitochondrialny.
Mt-Eva,
czyli ta, której potomstwo się rozgałęziło i istnieje do dziś,
żyła około 177,000
lat temu.
(oznacza to, że rozgałęzienia potomstwa jej poprzedniczek, także
rówieśniczki żyjącego około 200-280 tysięcy lat temu Y-Adama,
czyli właściwej mt-Ewy, nie dożyły do dziś.
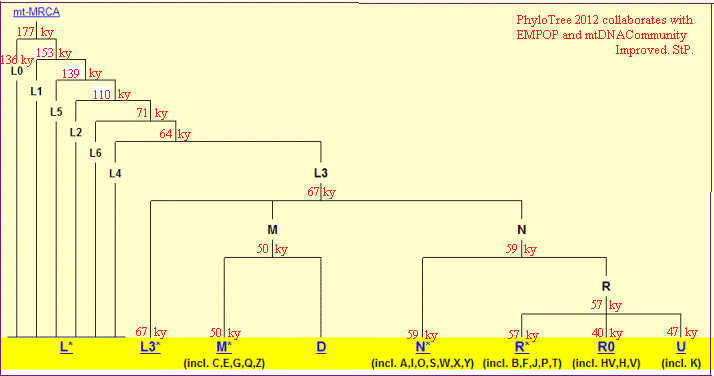
(Uwaga, niektóre daty haplogrup wymagają aktualizacji)
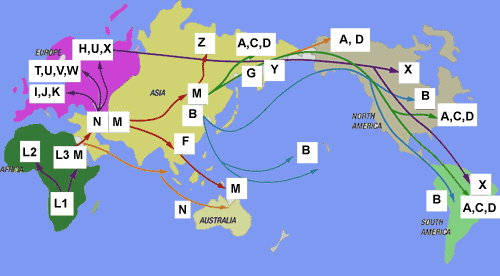
W
Wikipedii mapa mutacji i
migracji haplogrup mtDNA od mitochondrialnej Ewy z Afryki na
resztę świata.
Grupy europejskie
Powstała
na Bliskim Wschodzie haplogrupa mtN i
liczne jej podgrupy znajdują się, w przeciwieństwie do
haplogrupy mtM i jej podgrup, głównie w rejonie Kaukazu, Azji
Mniejszej i w Europie; zapewne one uczestniczyły
w powstaniu ludzkiej rasy kaukaskiej, europoidalnej, białej.
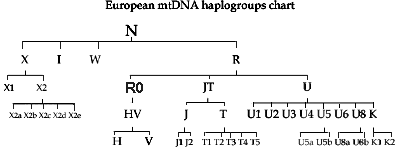
Natomiast haplogrupa
mtM i jej podgrupy -
upowszechniły na pozostałych kontynentach
(poza Afryką).
Na poniższej,
ogólnoświatowej mapie
brak wytworzonej z haplogrupy N i w jej regionie, a niedawno odkrytej, haplogrup mtR
(zob. wyżej!), od której
wywodzą się prawie wszystkie podgrupy europejskie, a zwłaszcza
poprzez mtRO i mtHV ważna europejska haplogrupa
H. Obejmuje ona w ponad 20 podgrupach około 40% ludności
europejskiej i polskiej. O obecności haplogrup mitochondrialnych
w rejonie Dunajca, zob.
W moim
rodzie podgrupą mtHV-1 wyróżniała się
Katarzyna Kuras
Szlagowa z Łukowicy. Popularną w Europie podgrupę
mtH miała
Anastazja Wilk Gurgulowa ze Stronia. Natomiast podgrupą
mtH-14a odznaczała się
Katarzyna Iwan Szlagowa z Łukowicy. Z haplogrupy
mtRO pochodzi m.in. podgrupa mtU, bardzo stara w Europie.
Właśnie grupą mtU3 wyróżniała się w moim rodzie
Anna Legutko
Janikowa, rodem z Jasiennej i Łazów Biegonickich.
MtDNA
przekazywane jest zarówno synom, jak i córkom, ale tylko przez matki.
Dlatego służą do rozpoznania tylko matczynego
pochodzenia, zarówno mężczyzn, jak i kobiet; podczas gdy Y-DNA
służy poznaniu tylko ojcowskiego pochodzenia, i tylko mężczyzn.
MtDNA ułatwiają też rozpoznanie tras
dawnych migracji rodów i plemion, choć nie tak wyraziście, jak męskie Y-DNA; mężczyźni
bowiem ściślej tkwili w swoich pierwotnych grupach rodowych i
plemiennych.
Np. mtDNA-H5 i H7, stwierdzane
w rejonie Kaukazu, wzdłuż Dunaju i pod Alpami, migrowało z męską
Y-hg. R1b. Podobnie mt-U3, stwierdzane wokół Morza Czarnego oraz
mt-J, rozpowszechnione pierwotnie w Azji Środkowej i wokół mórz
Kaspijskiego i Czarnego. Haplogrupie Y-R1a towarzyszyło
głównie mt-U4, bardziej popularne w rejonie Tadżykistanu i
Pakistanu, w Azji Środkowej oraz w Europie Wschodniej. Pochodząca z Afryki północno-wschodniej mt-T, stwierdzana
jest w
rejonie Morza Egejskiego i Europie Wschodniej; z
populacjami Y-R1a można łączyć też mt-U3 i mt-J (towarzyszące
jednak głownie Y-hg. R1b). Mt-H13 (i mniej H2) łączy Kaukaz i
Sardynię, podobnie jak męska Y-G2a. Mt-H1, H3, V i U5 zdradzają
paleolityczne i mezolityczne towarzystwo łowców-zbieraczy (Y-hg.
I); także mt-J1 występuje przy germańskich populacjach hg. I.
Mt-K1a towarzyszy głównie męskim J, T i E1b1b.
Ponieważ mitochondrialne DNA łatwiej niż Y-DNA
trwają niezniszczalnie w szczątkach kopalnych, mogą ułatwić
identyfikację oraz stwierdzenie ciągłości między
człowiekiem paleolitycznym a dzisiejszym, np. człowiek z Kostienek (nad Donem w Rosji), z około 30.000 lat temu,
odznaczał się haplogrupą U2; mtDNA kromaniońskiego człowieka z
Paglicci Cave (Włochy), z około 28.000 lat temu, ujawniło
jednoznaczną różnicę względem neandertalczyka, a tożsamość
z dzisiejszymi typami HV lub U.
* * *
* *
Zob.
opracowanie datowania mitochondrialnych haplogrup:
Zob.
Soares et al. 2009, datowanie:
Supplemental data: An Improved Human Mitochondrial Molecular Clocka
Nowe ustawienie drzewa mtDNA w
odniesieniu haplogrup do pierwszej mitochondrialnej matki,
mt-Ewy, z haplogrupą RSRS (177.000 lat
temu), zamiast dotychczasowego odniesienia CRS (Cambridge).
Spis haplogrup i datowanie ich wieku
(time estimate - years):
Behar et al. 2012,
http://www.sciencedirect.com/science/article/pii/S0002929712001462
Tamże, Suplement: Document S1.
Figures S1 and S2 and Tables S1–S6.
Pełne aktualne drzewo
genealogiczne mtDNA w ośmiu częściach:
http://www.phylotree.org/tree/main.htm
Fu et al. 2013: A revised timescale for human evolution based
on ancient mitochondrial genomes
http://ac.els-cdn.com/S0960982213002157/1-s2.0-S0960982213002157-main.pdf?_tid=00a2a2d8-3a89-11e5-962d-00000aab0f26&acdnat=1438679760_ba8d2d7655913df101be8e1fcdb79cc0
Drzewo genealogiczne i mapa archeologicznych znalezisk
Denisowczyka, Neandertalczyka i człowieka współczesnego.
* * *
* *
Ilustracją obecności poszczególnych haplogrup mtDNA
na polskiej ziemi są rezultaty analiz w ramach polskiego
programu/projektu
Family Tree DNA, który administruje L. Mayka (tablica
może otwierać się
powoli, podobnie jak tam dołączona mapa).
 |
Polskie mtDNA w testach FTDNA
Kolory pin. wskazują na haplogrupy mtDNA.
Oto przybliżone ilości w poszczególnych haplogrupach:
C-10, D-3, H-200, I-10, J-60, K-60, L-10,
N-20, R-10, T-50, U-70, V-20, W-20, X-20. Razem 563. |
W sumie,
inaczej niż ojcowski rodowód, matczyny rodowód Polek i Polaków
przeważnie niewiele
różni się od pozostałych krajów Europy (zob.
na tablicy Eupedii). Wyraźne różnice zachodzą natomiast w
stosunku do ludności krajów Azji i Afryki,
jak wskazuje mapa migracji.
Genetycy starają
się też ustalić haplogrupy wyróżniające populacje słowiańskie w
Europie; zob. np.:
1) Grzybowski et al. 2007:
Complex interactions of the Eastern and Western Slavic
populations with other European groups as revealed by
mitochondrial DNA analysis.
2) Malyarchuk et al. 2008:
Mitochondrial DNA Phylogeny in Eastern and Western Slavs.
3) Malyarchuk
et al. 2010:
The Peopling of Europe from the Mitochondrial Haplogroup U5
Perspective
Tabela mtDNA dotąd wykrytych w
archeologicznych szczątkach ludzkich
w znanych kulturach archeologicznych, głównie europejskich
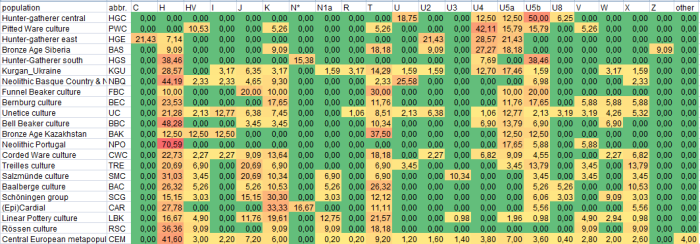
Zob. praktyczne zastosowanie
genealogii mtDNA na rodzinnej stronie:
Szlagowie.
UWAGI
a) Dlaczego tak szczegółowo, jak na internetowe zwyczaje, z
podaniem podstawowej terminologii i literatury, przedstawiam ten temat?
Dlatego, że dla mnie i wielu z nas bardzo ważne jest pytanie: Jaka jest nasza
genealogia - skąd jesteśmy,
co nas łączy lub dzieli, jaka jest nasza przeszłość, dlaczego nie możemy się
jej wyrzekać, a wreszcie dlaczego nie powinniśmy pogardzać swoją słowiańskością
i polskością!
Genealogia Y-chromosomowa (Y-DNA) bardzo służy historii, poznaniu genezy pierwotnych
ludzkich plemion, większych populacji i narodów oraz ich przemieszczaniu się po
Ziemi. Ułatwia poznanie wydzielonych męskich rodów i w ogóle rodzin, które nas
wydały na świat, do których należymy i wobec których mamy obowiązki - nie tylko
wdzięczności! Zdecydowanie pomaga określić twórców archeologicznych kultur,
ujawnianych w pracach wykopaliskowych.
Ma to szczególne
znaczenie prestiżowe dla Słowian. Rzekome późne wyłonienie się etnosu
słowiańskiego (przypominam: gr. "etnos" to rój, stado, plemię, a nie
sama kultura!) bywało nieprzychylnie dla
Słowian interpretowane. Do tego przyczyniło sie także i nazewnictwo. Mianowicie,
ponieważ we wczesnym średniowieczu Słowianie bywali przez kupców arabskich i
żydowskich sprzedawani na rynkach Europy jako niewolnicy, od etnonimu Słowianin,
łac. Sclavus, utworzono około X w. łacińskie słowo "sclavus", oznaczające
niewolnika, sługę, a także w ogóle człowieka gnuśnego. I to określenie w rozmaitej formie (np. sclave slave, esclave, slav...) przedostało się do innych języków zachodnich.
Niekiedy niestety wpływa ono na opinię o ludach słowiańskich i na sposób
traktowania Słowian przez zachodnich Europejczyków.
b)
Rozpoznawanie swojego cennego rodowodu w ojcowskim Y-DNA czy macierzyńskim mtDNA i wydzielanie
krewniaczych haplogrup ludzkich nie oznacza tworzenia jakiegoś nowego rasizmu,
tym razem genetycznego, na
miejsce dotychczasowego, opartego zasadniczo tylko na zewnętrznym wyglądzie człowieka. Genealogia Y-DNA czy mt-DNA umożliwia
zidentyfikowanie i umieszczenie osób czy rodu na ojcowskim czy
macierzyńskim drzewie genealogicznym ludzkości i ustalenie czasu wyodrębnienia
się, co w sumie już daje wielkie poczucie zakorzenia i wspólnoty, ale to
dopiero ułamek
prawdy o pochodzeniu konkretnej osoby.
Bowiem oprócz niewielkiego
chromosomu płciowego Y-DNA lub mitochondrialnego mtDNA posiadamy daleko większe 22
inne pary chromosomów, tzw. autosomalnych, i ponad trzydzieści tysięcy innych genów kodujących. Przekazywane są one zarówno
przez ojca, jak i przez matkę, i zarówno synom, jak i córkom. One kształtują
podstawowe funkcje organizmu, a także kolor skóry, oczu, czy inne nawet bardzo
drugorzędne drobiazgi. W ten sposób każdy
dzisiejszy człowiek ma pozostałe chromosomy i geny nie tylko od około 8.000
swoich dotychczasowych przodków w linii prostej aż do Adama i Ewy, ale także,
jak obliczyli antropologowie, od około 80.000 innych przodków z bardzo
rozbudowanych linii bocznych.
Świadomość tego, że nosimy w sobie materiał
genetyczny tylu, z rozmaitych haplogrup i ras przodków, powinna - niezależnie od
ustalonej haplogrupy Y-DNA lub mt-DNA - chronić od rozmaitych ideologii
rasistowskich i pozytywnie wpływać na poczucie braterstwa z
wszystkimi istotami ludzkimi. Genetyka antropologiczna już poszukuje w
autosomach, czyli chromosomach pozapłciowych, owych mutacji "rasowych", czyli
kształtujących odrębny wygląd i cechy fizyczne poszczególnych
grup ludzkich oraz ustala na mapach świata trasy
"migracji" dziedziczonych genów, tak jak już od dawna czyni to genetyka medyczna, identyfikująca obciążone chorobami
dziedzicznymi osoby i populacje.
c) Ponieważ wraz z odkrywaniem
nowych mutacji w SNP zmienia się klasyfikacja haplogrup na
drzewie genealogicznym,
jak ostatnio zmieniono R1a1 na R1a1a i R1a1a7 i inne, a nadto
stale napływają nowe etniczne i geograficzne ustalenia co do
poszczególnych populacji, dlatego co pewien czas może
zajść potrzeba dokonania poprawek w tym artykule; mogą też
powstać niezgodności z poprzednimi tekstami i cytowanymi
pracami. Trzeba też śledzić zmiany w terminologii, nazewnictwa i
symboli haplogrup. Np. dla uproszczenia umownie podaje się
krótszą nazwę haplogrupy ojcowskiej
zamiast wszystkich synowskich, np. ojcowską R1b zamiast
kilkudziesięciu haplogrup potomnych, ze skomplikowanymi ich
określeniami; albo R1b1b2 dla określenia
wszystkich europejskich jej podgrup. Niekiedy podaje się tylko
krótsze określenie mutacji, zamiast dłuższego określenia
odpowiedniej haplogrupy - miejsca na drzewie genealogicznym, np. R-M458 zamiast R1a1a7 albo R-U152
zamiast R1b1b2a1a2d - alpejskich i italskich Celtów. Nadto w
literaturze, niekiedy niekonsekwentnie, dodaje się gwiazdkę, np.
R1a*, dla określenia, że nie bierze się pod uwagę jej podgrup.
* * *
|
P.S. W Dziejach Apostolskich 16,9 i
nast. jest fragment o św. Pawle Apostole:
"/W Troadzie/ w nocy miał Paweł
widzenie: jakiś Macedończyk stanął przed nim i błagał go:
'Przepraw się do
Macedonii i pomóż nam' ".
Dalej Dzieje Apostolskie podają, jak Paweł
zrealizował prośbę Macedończyka
i jaki był skutek jego apostolskiego działania.
Zgodnie z tym, co nam podsuwa
genetyka (jak wyżej), wolno nam przypuszczać,
że tym Macedończykiem był jakiś
Słowianin, potomek tych, którzy byli tam już w co
najmniej w szóstym tysiącleciu przed Chr., symboliczny typ naszego praojca w linii prostej.
Nawrócona tam
przez Apostoła Lidia była podobno natchnieniem dla H.
Sienkiewicza
do wykreowania postaci słowiańskiej Ligii, bohaterki "Qvo vadis".
Jakieś wyczucie? |
Jak zbadać swoje Y-DNA i mtDNA
|
Najlepiej dokonać badań w FamilyTreeDNA (FTDNA) lub
innej firmie, zachowującej pewne standardy.
Proponuję zbadać Y-DNA - 67 merkerów, bo to teraz
standard i zarazem mtDNA w rejonach HVR1 i HVR2, bo to
też standard.
Tym, którzy testują w FTDNA, proponuję od razu
przyłączyć się do PolishProject FamilyTreeDNA (będzie
może trochę taniej i jest jakaś "opieka" administratora
polskiego pochodzenia. Lawrence Mayka jednak nie zna
polskiego!).
1) FTDNA Polish Project.
Otworzyć:
http://www.familytreedna.com/group-join.aspx?Group=Polish
2) Wybrać rodzaj testu (aktualne ceny tam podano)
* Y-DNA37, $119
* Y-DNA67, $189
* Y-DNA111, $289
* mtFullSequence, $169
Dla tych, którzy już mają podstawowy test:
Test całego Y-DNA (w regionie NRY)
Big Y-DNA test, $495 (do 01.12.2013) lub #695 po
01.12.2013
zob.
Big Y - FAQ
3) Wypełnić formularz osobowy:
- prefix: wybrać mr.
- imię
- nazwisko (last name)
- adres zamieszkania (miejscowość, ulica, nr. domu - dla
przesłania kitu)
- miejsce poczty (city)
- kod pocztowy
- kraj (country)
- telefon
- e-mail (dla powiadomień i korespondencji)
- powtórzyć e-mail
To wszystko w formularzu.
4) Na kolejnej stronie kliknąć-wypełnić sposób zapłaty,
np. kartą kredytową lub na fakturę (Invoice to na
fakturę, także jeśli ktoś płaci
przez PayPal przed lub po przysłaniu faktury).
Zatwierdzić i koniec.
5)
Po godzinach lub dniu przyślą podziękowanie, numer
zestawu badań "Kit" i hasło oraz podadzą kwotę
zapłaty.
Teraz można wysłać zapłatę.
Potem przyślą w kopercie trzy "szczoteczki" do zrobienia
trzech wymazów z wewnętrznej strony policzka i trzy
pojemniczki z płynem na te próbki.
Trzeba zrobić wymaz/próbki zgodnie z instrukcją, sanitarnie
(czyli bez dotykania palcami "szczoteczki"),
potem nacisnąć na patyk wystający z rurki trzonka tej
"szczoteczki", aby sama szczoteczka odpadła do
przysłanego pojemniczka (z płynem), zatkać i odesłać w
otrzymanej kopercie.
Czekać na wynik około miesiąca. W tym czasie można
przestudiować pytania w FAQ.
6)
Już po otrzymaniu nr kit i hasła można urządzić swoją
stronę internetową My FtDNA Home:
Po wpisaniu nr kit i hasła otworzy się strona: MY FTDNA HOME
Pod MY MAPS kliknąć: Plot ancestral locations.
Wpisać tam imiona i nazwiska najdawniejszych przodków Y
i mt, ich daty (rok) i miejscowość ich urodzenia lub -
jeśli nieznane urodzenie - miejsce przebywania.
Google może automatycznie podać współrzędne geograficzne
tej miejscowości; można akceptować lub można zmienić, a
korzystając z przybliżeń - można dokładnie doprecyzować
lokalizacje obydwu pinesek.
Przysłany wynik publikuje się automatycznie w
odpowiednich miejscach na stronie MY FTDNA, a sam
testowany swoich przodków umieszcza na mapie, w bazie
YSearch lub w innych miejscach - według propozycji FTDNA.
7)
Można też osobiście przekazać dane na rozmaite
wykresy drzewa i mapy genealogiczne oraz zestawienia
zbiorcze zwane projektami - a to dla celów
rodzinnych (genealogicznych) lub naukowych.
Do nich należą np.:
a) Polish FamilyTree DNA Project, zestawienie
haplotypów wszystkich haplogrup, związanych z Polską w
jej dawnych granicach); administrator Lawrence Mayka
http://www.familytreedna.com/public/polish/default.aspx
b) R1a and Subclades Project (zestawienie wszystkich
haplotypów haplogrupy R1a i podgrup na świecie);
Projekt międzynarodowy dla wszystkich R1a. Administrator Łukasz Łapiński
http://www.familytreedna.com/public/R1a/default.aspx
Dla każdej haplogrupy istnieją osobne projekty FTDNA.
Aby dołączyć się do jakiegoś projektu, wystarczy
skorzystać z odpowiedniego linku na swojej stronie My
FTDNA Home.
|
Ilustracją, jaki może być użytek naukowy z każdego wykonanego
testu, jest treść strony:
"Genetyka odsłania
tajemnice: Pochodzenie ludności w dorzeczu Dunajca i jego
sąsiedztwie"
Adres: ks. Stanisław Pietrzak,
Tropie 6,
33-316 Rożnów
|
Zadzwoń,
przekaż uwagę lub informację: |
tel. 018.449 41 81
lub 601 918 322 |
(można dzwonić
do 2100)
E-mail:
pietrzakstan@poczta.onet.pl
Wszelkie uwagi dla mnie proszę
przekazać meilowemu gołąbkowi:

Dziękuję,
już dziś przekażę Adresatowi
|
DZIĘKUJĘ!
JESTEŚ MIŁYM |
 |
GOŚCIEM
NASZEJ WITRYNY!
|
Do strony Pietrzakowie. Genealogia.
Pietrzak
Do strony Kurczabowie. Genealogia.
Kurczab. Bołoz
Do strony Janikowie. Genealogia. Janik
Do strony Szlagowie. Genealogia.
Szlaga/Ślaga
Powrót do Strony
głównej witryny: Parafia TROPIE
Str.pryw.-genealog.
PIETRZAKOWIE.Pietrzak,
JANIKOWIE.Janik,
FIUTOWIE,
SZLAGOWIE Nadto:
Gierałt.GIERAŁTOWIE
|