|
Osiem tysięcy praojców R1a1a1-M417* i H14a |
Skąd
pochodzą Rozmaitości genealogii genet. (część 2) |
Ostatnia
aktualizacja |
†
WYBIERZ RÓD:
|
nadto: GIERAŁTOWIE |
POLACY, SŁOWIANIE... |
Strony genealogii genetycznej na naszej witrynie
1.
Genealogia Y i mtDNA, cz.1, haplogrupy R1a, R1b
2.
Geneza języków
indoeuropejskich i początek Słowian
3.
Haplogrupy nieindoeuropejskie
Y-DNA. Haplogrupy mitochondrialne mtDNA
Jak zbadać swoje Y-DNA i
mtDNA
Pochodzenie ludności w dorzeczu Dunajca i jego sąsiedztwie * Inne wybrane regiony
Ilustracje R1a1a: drzewa, mapy, filogeneza
Skąd, Europejczyku, jak i kiedy tu przybyłeś?
Nowiny i linki do naukowych publikacji
Rozmaitości genealogii genetycznej i dyskusja [cz.1] [cz.2]
Dyskusja o mutacjach STR. Faktyczny czas mutacji. TABLICE
Y-Adam, praojciec wszystkich współczesnych ludzi [1]
[2] *
Fundusz RRA00
[1]
[2]
Kreacjonizm
ewolucjonistyczny
The evolutionistic
creationism
ROZMAITOŚCI
GENEALOGII GENETYCZNEJ i DYSKUSJA
[część
2]
[1] 07.10.2010.
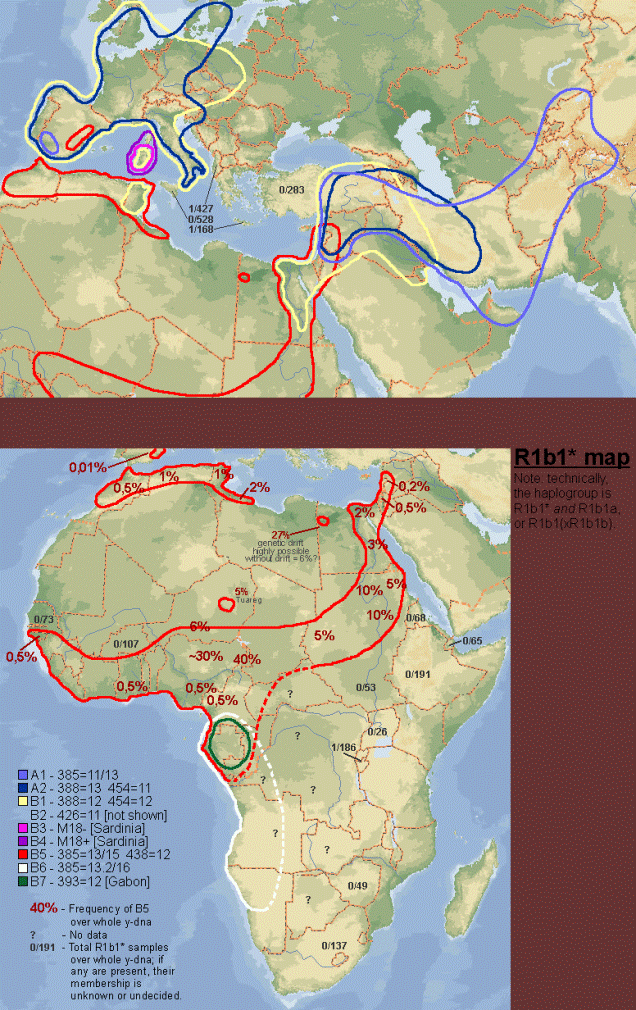
Mapa rozwoju wczesnej haplogrupy: R1b1*
w Azji Środkowej, na Bliskim Wschodzie, Europie (w tym
Sardynia) i w Afryce
(głos w dyskusji na temat haplogrupy R1b1b2).
|
http://www.worldfamilies.net/forum/index.php?topic=9360.0
|
kolejność |
|
|
1. niebieski |
[2] 07.10.2010.
Papierowe publikowanie prac genealogii
genetycznej. Czy potrzebne?
Jan L. napisał: Witam
serdecznie, od dłuższego czasu z ogromnym zainteresowaniem
odwiedzam Pańską stronę. Ma Pan dar interesującego i klarownego
przekazywania wiadomości, przynajmniej na ten temat :).
Czy nie myślał Pan o książkowym wydaniu, tej bardzo już
obszernej wiedzy? Zdaje sobie sprawę, że nasze wiadomości na ten
temat bardzo dynamicznie poszerzają się, co z pewnością
znakomicie utrudnia taki projekt. Jednak popularyzatorskie
znaczenie takiej pozycji byłoby trudne do przecenienia. Mimo
sporej dostępności internetu, słowo drukowane ma wciąż większy
zasięg, a z pewnością ma dużo większe znaczenie. Dziękuje za
Pańską stronę, pozdrawiam Jan L.
Odpowiadam.
Aby publikować prace genealogii
genetycznej, mające charakter podsumowania, trzeba mieć dużo
beztroski. Bowiem każdego prawie tygodnia pojawiają się różne
publikacje, często sprzeczne z sobą, na których ocenę trzeba
poczekać do... kolejnych kontrowersyjnych publikacji.
Relacjonowanie rozmaitych poglądów bez ich własnej oceny ma
ograniczoną wartość. Zaś dokonanie ocen, wybór najlepszej
alternatywy wymaga czasu i nowych odkryć. Odkrycie jakiejś
jednej ważnej mutacji może zmienić całe drzewo filogenetyczne
tej czy innej mutacji, czyniąc poprzednie publikacje niemal
bezużytecznymi. To czyni pracę autora bardzo stresogenną.
Nadal czekamy na jednolita stawkę mutacji na haplotyp na
pokolenie i dopasowanie do do niej wszystkich prac. Ostatnia
praca
Ballatyne et al. 2010, podająca stawki mutacji w parach
ojciec-syn jest niezmiernie ważna. Jednak pozostaje otwarty
problem liczenia mutacji, na który genealodzy nie bardzo
zwracają uwagi. Mianowicie wraz ze wzrostem liczby mutacji w
haplotypie o tyle maleje szansa na taką następną i o tyle
wzrasta szansa na mutację zwrotną, przeważnie wymazującą mutację
poprzednią, czyniącą siebie i poprzednią niezauważalnymi (zob.).
Stąd pozostaje jakieś zapotrzebowanie na stawki "ewolucyjne"
Żywotowskiego, sprzeczne ze stawkami przyjętymi w
genealogii. Itepe, itede! Jak je pogodzić z sobą?
Dlatego - jak dotąd - najlepszą
publikacją dla genealogii genetycznej jest strona internetowa,
którą codziennie można aktualizować. Zresztą, strony internetowe
powoli urastają do rangi właściwego miejsca publikacji prac
naukowych. Oczywiście pod określonymi warunkami. Te warunki
kiedyś będą ustalone - np. jednolity i stabilny rejestr i
datowanie aktualizacji.
[3] 07.10.2010.
Migracyjny szlak
haplogrupy R1b1b2 do Europy
(krytyczna analiza artykułu
dokonana przez A. Klosowa)
P. Balaresque et al. "Przewaga neolitycznego początku
europejskich linii ojcowskich"
Миграционный путь гаплогруппы R1b1b2 в Европу (1). Критический
разбор статьи
“A Predominantly Neolithic Origin for European Paternal Lineages”,
Patricia Balaresque et al,
PLoS Biology, vol. 8, January 2010, on-line publication. А.А.
Клёсов, str. 896
Balaresque stwierdza, że dotychczasowe badania (oczywiście
badania genetyki populacyjnej) wskazywały na paleolityczne
pochodzenie tej haplogrupy w Europie. Ale w tym badaniu
geograficznego rozmieszczenia różnorodności STR najlepiej je
wytłumaczyć rozprzestrzenianiem się z jednego źródła na Bliskim
Wschodzie poprzez Anatolię w okresie neolitu. Porównawcze
badanie pochodzenia innych haplogrup też wskazuje na czas
ekspansji neolitu; w przeciwieństwie do większości linii
matczynych, pochodzących od łowców-zbieraczy, sugerujących tylko
przejście kulturowe do nowego systemu gospodarowania. W ten
sposób Europa staje się doskonałym przykładem zmian
technologicznych i kulturowych, związanych z rozwojem męskich
linii Y-DNA i wyjątkowej roli mężczyzn w okresie przejściowym.
Wielu genealogów genetycznych
(np. na forum RootsWeb) tę pracę Balaresque et al. przyjęło
pozytywnie, bo przyznała ona konkretną rolę wielkiej haplogrupy
R1b1b2 po jej przybyciu do Europy. Nadto bezkrytycznie zauważyli
tu jakoby rozwiązanie zagadki upowszechnienia się neolitu na
kontynencie europejskim - czyim było ono dziełem. Pozytywnie też
odebrali oczekiwane odejście panującego w kręgu genetyki
populacyjnej "współczynnika Żywotowskiego", zawyżającego czas
mutacji blisko trzykrotnie i o tyle przesuwający dzieje
populacji w daleka przeszłość.
A co na
to A. Klyosov?
Biorąc pod uwagę głośne
nazwiska autorów tej publikacji Balaresque i kol., uznał
charakter pracy, metodologię i sposób dowodzenia za
"przerażające, bez przesady" (ужасные, без преувеличения).
Po
pierwsze, autorzy, zebrawszy z różnych regionów Europy
ogromny i cenny zestaw haplotypów R1b1b (M269) w liczbie 2574,
jakby nawet nie podejrzewali, że w tym zestawie znajduje się
wiele kolejnych i daleko późniejszych mutacji i, wskazujących na
nowe, późniejsze populacje, mające też może oddzielne ścieżki
migracji.
Po
drugie, dzisiejszym regionalnym skupiskom haplotypów
autorzy bez zastrzeżeń przypisują starożytne neolityczne
kultury, byleby łączyła ich niewielka odległość geograficzna –
do 150 km. W ten sposób lokalnym skupiskom haplotypów ustalono
daty owych kultur, ustalone radiowęglowo, np. w zachodniej
Turcji – 9000 lat, w centrum 9800 lat, a na wschodzie 10 200 lat
temu. Nadto przypisali regionalne kultury haplogrupie R1b1b2,
choć mogli uwzględnić także inne, np. G, J2, E, R1a lub inne,
np. wymarłe. Podobnie uczyniono w innych regionach Europy, np.
na Płw. Pirenejskim, gdzie według badań Klosowa haplogrupa G
liczy przecież 7600 lat, J1 – 9450, J2 – 7000 a K – 9100 lat.
Po
trzecie, autorzy nie podają stosowanych stawek mutacji,
a z wyników widać, że posługiwali się jakimiś zawyżonymi. Podają
też dziwnie szeroki zakres błędu, np. 10490-4423, tak aby
umieścić w nim dowolną kulturę o dowolnym datowaniu.
Po
czwarte, autorzy wskazali na rozprzestrzenienie się
populacji z Bliskiego Wschodu przez Anatolię z ” jednego
źródła”, ale nie dali żadnego uzasadnienia; zamilczeli inne
znane możliwości; a choć cytowali liczbę haplotypów w Rosji,
zamilczeli ich znane i wczesne datowanie. Dlatego łatwiej mogli
podać datę pojawienia się populacji R1b1b2 w Europie 8-7 tysięcy
lat temu, czyli na czas pojawienia się neolitu (zamiast 5-4
tysięcy lat temu).
Recenzujący przy okazji przypomina znane i opublikowane
własne ustalenia.
1)
W Turcji R1b1b2 pojawiło się nie wcześniej niż 6000 lat temu, a
na kontynencie europejskim nie wcześniej niż 5000 lat temu, a
dokładniej nie wcześniej niż 4800-4600 lat temu. W tym samym
czasie pojawiło się i na Płw. Pirenejskim, około 4800-4600 lat
temu, z przejściem w głąb Europy 4500-4200 lat temu, i dalej po
Europie 3700 lat temu.
2)
Rzecz w tym, że R1b1b2 przyszło do Europy nie jednym
szlakiem i w jednym czasie, a różnymi szlakami i różnymi
subkladami (podgrupami). Jedne z południa na Płw. Pirenejski,
inne z Małej Azji na Bałkany i przez Italię, inne wreszcie z
Ruskiej Równiny (gdzie wspólny przodek liczy około 7000 lat) na
zachód (być może także szlakiem północnym, nadbałtyckim). Z
Ruskiej Równiny około 6000 lat temu migrowali też na Kaukaz,
skąd do Anatolii i na Bliski Wschód (tu mogli pojawić się też
bezpośrednio z Południowej Syberii).
3)
Na Ruską Równinę R1b1b2 przyszło z Uralu i Azji
Centralnej , dokąd z kolei razem z R1b1b1 zapewne z Południowej
Syberii. Wspólny przodek ich i europejskich haplotypów żył 16
000 lat temu. Dotąd R1b i podgrup nie znaleziono w Południowej
Syberii, ale należy ich tam się spodziewać ze względu
nieobecność tam pokrewnych haplogrup Q i R1a, które tam są
zarazem najstarsze. Ta druga, czyli R1a, w przylegających
prowincjach Chin (Hui, Sala, Bonan i Dongxiang) spotykana jest w
częstotliwości odpowiednio 18%, 21%, 25% i 32% ! (Bittles et al.
2007).
Nadto tu w trzech ostatnich prowincjach spotkano ojcowską
haplogrupę P w częstotliwości 3,2%, 5,6% i 9,2%.
4) I to
bardzo ważne. W kilku północno-zachodnich
prowincjach/narodowościach Chin (Salar, Tu i Bao’an) znaleziono
nielicznych (10/503) przedstawicieli R1b* (M343) z datą
wspólnego przodka 11 325 lat temu. W zestawieniu z atlantyckim
haplotypem modalnym daje im 21 000 lat do wspólnego przodka
(str. 901).
[4] 14.10.2010.
Chronologia i migracje klanu R1b i podgrup w Azji, Europie i Pn. Afryce
Natalie Myres - Hua Zhong - Anatolij Klosow (AKlyosov)
W ostatnim czasie ukazały się cztery ważne prace na temat populacji (klanu) haplogrupy R1b i głównych jej podgrup, ważnych dla znacznego obszaru kuli ziemskiej, a zwłaszcza Europy.
1) N. Myres et al. 2010,
A major Y-chromosome haplogroup R1b Holocene era founder effect
in Central and Western Europe.
2) Krytyczna analiza artykułu:
A. Klosow, Migracjonnyj putj gaplogruppy R1b1b2 b Europu i rassedenije w
Europe ( w j. rosyjskim),
w: Wiestnik Rossijskoj Akademii DNK-genealogii, tom 3, nr 10,
2010, s. 1652-1675.
3) H. Zhong et al. 2010,
Extended Y-chromosome investigation suggest post-Glacial
migrations of modern humans into East Asia via the northern
route.
4) Krytyczna analiza artykułu:
A. Klosow, Gaplogruppa R1b1 i jeje subklady w Azji,
w: Wiestnik Rossijskoj Akademii DNK-genealogii, tom 3, nr 10,
2010, s. 1676-1695.
Obydwa artykuły A. Klosowa są nie tylko krytyczną analizą prac N. Myres i H. Zhong, lecz także nowym opracowaniem opublikowanych przez tych autorów haplotypów Y-DNA.
To, co najwięcej należy kwestionować w pracach obydwu zespołów "genetyków populacyjnych", to próba rozwiązywania problemów etnogenezy neolitycznych kultur archeologicznych przy pomocy materiału Y-DNA zupełnie nieprawidłowo datowanego, tak zwaną metodą Żywotowskiego. By można ją bezpiecznie zastosować w jakimś przypadku, nie wystarczy, jak czynią niektórzy, jej wyniki dzielić po prostu przez trzy, gdyż oparta na proporcjach różnorodności, czyli wielości mutacji w puli genowej bez grupowania haplotypów według przynależności do wspólnych przodków, daje wyniki datowania nierównomiernie, od 2 do 4 razy zawyżone.
Dzięki takiemu datowaniu Mures et al. mogli dojść do wniosku, że populacji italo-celtyckiej haplogrupy R-P312/S116 (której początek w Europie inni autorzy datują na 4500 lat temu) można przypisać rozprzestrzenienie kultury ceramiki liniowej rytej około 10000 (sic!; faktycznie 7500 6500) lat temu. Nie wyklucza się udziału w tym i starszego klanu, R-L23 (choć faktycznie to prawidłowo byłoby tę kulturę przypisać klanom hg. I i R1a1a.
Autorzy uznali, że R-L51/M412 (ojcowski dla R-P312) jest tym, który oddziela środkowo i zachodnioeuropejskie populacje R1b od wschodnioeuropejskich i azjatyckich. Wskazali na dwa ośrodki koncentracji klanu P312/S116: nad Górnym Dunajem (rejon Monachium) i w rejonie Paryża (choć faktycznie zupełnie zignorowali szczególne miejsce koncentracji, zwłaszcza R-P312 na Półwyspie Iberyjskim).
Myres et al. uznali, jakoby przybycie klanu R1b1b2 z Azji do Europy korespondowało z przybyciem i rozprzestrzenieniem sie rolnictwa w Europie. Właściwie bez dowodu, prócz tej fatalnej datacji. Powołali sie zasadniczo na omówioną tu wyżej pracę Balaresque (2010), która również ma poważne usterki rzeczowe (brak dowodów rzeczowych) i co do datowania.
Praca Myres i kolektywu ma jednak zasadniczą wartość. Prezentuje ona wielki zestaw 2043 haplotypów (choć krótkich) haplogrupy R1b1b2 i jej podgrup ze 118 krajów Azji Zachodniej i Europy, oraz mapy ich rozprzestrzenienia się w tych regionach. Ułatwiają one opracowanie daleko idących wniosków. Klosow te wnioski, częściowo już znane z jego poprzednich licznych publikacji, zebrał w Wiestniku t.3, nr 10 (październik 2010) na stronicach 1656-1657. Powtórzone one zostały tamże, w drugim artykule, na str. 1680-1681.
Podstawą tych wniosków jest bowiem także druga, równoległa praca, H. Zhong et al. 2010, dostarczająca znaczną ilość, bo aż 3826 haplotypów R1b i innych (w tym ze 116 populacji Chin i jednej z Południowej Korei) i innych krajów Azji, z uwzględnieniem ich geograficznego rozprzestrzenienia. Autorzy i tym razem posłużyli się błędną datacją Żywotowskiego i wyprowadzili kilka nieprawidłowych wniosków, jak choćby ten, że populacje haplogrupy Q i R migrowały do Wschodniej Azji szlakiem północnym około 18000 lat temu (epoka szczytowego zlodowacenia!), choć skądinąd już wiadomo, że haplogrupa Q była we wschodniej Syberii już około 46 tysięcy lat temu, dzięki czemu mogła ona pojawić się mniej więcej wtedy na kontynencie północnoamerykańskim. Nie powiodła się im też próba ustalenia migracji haplogrupy NOP jakimś południowym szlakiem na wschód. Autorzy zresztą nie przedstawili żadnej analizy prezentowanych w pracy haplotypów.
Jaka
rzeczywistość wyłania się z analizy tych, przeważnie krótkich, ale
jednak bogatych zestawów haplotypów Y-DNA Europy i
Azji? Rosyjska genealogia Y-DNA, która w przeciwieństwie do
europejskiej, zajmuje się także wcześniejszymi, azjatyckimi
etapami migracji populacji R1a i R1b, dopracowała się następującego
obrazu migracji klanu R1b.
Haplogrupa R1b1, jak i pokrewna jej R1a1 wyszła z Południowej Syberii i przyległych terytoriów Ałtaju i północnych Chin. Obie haplogrupy europoidalne i ich ojcowska haplogrupa P przybyły w rejon Ałtaju około 45-50 tysięcy lat temu z Niziny Wschodnioeuropejskiej (wg. Klosowa) lub raczej bezpośrednio od półwyspu Arabskiego czy Bliskiego Wschodu (wg. W. Ryżkowa). Tam haplogrupa P rozeszła się terytorialnie i genetycznie na haplogrupy Q i R. Nosiciele haplogrupy Q migrowali w kierunku Ameryki, jak wskazuje analiza radiowęglowa, około 46000 lat temu, co wyznacza zarazem datę rozejścia się P i Q w Południowej Syberii.
Haplogrupa R1b
formowała sie około 16000 lat temu w rejonie Ałtaju lub
sąsiedniej Kirgizji (Ryżkow natomiast,
analizujący paleokraniometrię, dla narodzin R1b wskazuje
rejon Płaskowyżu Irańskiego, po uprzednim przedlodowcowym
pobycie klanu R, R1a i preR1b na Rosyjskiej Nizinie - w Kostienkach i Sungirze i przedlodowcowej migracji na południe;
w okresie polodowcowym nastąpił powrót niektórych potomków na
północ, na Powoże i do Baszkirii).
Nosiciele tej haplogrupy R1b migrowali na zachód wzdłuż granicy
północnego Kazachstanu (uczestnicząc tam w tworzeniu kultury
botańskiej z późniejszym udomowieniem konia około 4500-3500
p.n.e). Około 11000 p.n.e pojawili się na terytorium
współczesnej Baszkirii, tworząc tam serogłazowską archeologiczną
kulturę (11000-9000 p.n.e)., następnie kulturę środkowowołżańką
(6000-5000 p.n.e.), samarską (5000 p.n.e.), sredniostogowską
(5500-3500), chwałyńską (5000-4000 p.n.e.), kamską (5000-3000
p.n.e.) - nazwane przedkurhanowymi.
Tam powstała haplogrupa R1b1b2-M269 i jej podgrupa L23. Z rejonu Baszkirii i Powołża jeden potomek tej populacji, z haplogrupą R-U106, udał się szlakiem północnym przez ziemie polskie w kierunku Niemiec.
Drugi, wyróżniający się mutacją L-23, potomek haplogrupy R1b1b2-M269 udał się przez Kaukaz do Anatolii (4000 p.n.e). Tak więc nie można jego rodowi przypisać anatolijskich kultur archeologicznych 8-7 tysiąclecia p.n.e. Klan ten otrzymał później, zapewne już w dolnym międzyrzeczu Tygrysu i Eufratu, etnonim "Sumerowie" (3500 p.n.e.) - twórcy wielkiej światowej cywilizacji w Mezopotamii. Migrowali oni później poprzez Egipt i północną Afrykę do wybrzeża Atlantyku (u Berberów około 3000 p.n.e.). Grupa R1b1, która wcześniej dołączyła się do nich około Palestyny lub Jordanii (gdzie ta haplogrupa żyje do dziś), w trakcie migracji odłączyła się i udała na południe, gdzie żyje do dziś w rejon Czadu i Kamerunu jako R1b1-V88, z TMRCA około 2600 p.n.e. Druga zaś część tej populacji przez Gibraltar przedostała sie do Europy na Półwysep Pirenejski (2800-2500 p.n.e), skąd z kulturą pucharów dzwonowatych przeszła dalej, do kontynentalnej Europy. Stało sie to pod naciskiem plemion Berberów z haplogrupą E1b1, które w końcu zupełnie przerzedzili na Pirenejach grupę R1b1b2 (i podgrupy L51 lub też L23), dlatego po przejściu tej "szyjki butelki" ta populacja datuje swój "początek" dopiero na 1700 p.n.e., ale już jako haplogrupa R-P312. To właśnie wspólny przodek Basków i większości innych populacji R1b1b2 Hiszpanii i Portugalii.
Dotychczasowy język, z którym haplogrupa R1b1b2-M przybyła do Europy był aglutacyjny, nieindoeuropejski, najprawdopodobniej z grupy sino-kaukaskiej (zwany prototurskim). Taki przetrwał do około początku lub połowy I tysiąclecia p.n.e. Taki język rozpoznawany jest u wielu plemion północnego Kaukazu, Sumerów, Basków i szeregu innych nieindoeuropejskich populacji.
Uzasadnienie. C.D.N.
[5] Human Y-chromosome short
tandem repeats: A tale of acculturation and migrations as
mechanisms for the diffusion of agriculture in the Balkan
Peninsula
American Journal of Physical Anthropology, Volume 142, Issue 3,
pages 380–390, July 2010
Support info:
http://onlinelibrary.wiley.com/doi/10.1002/ajpa.21235/suppinfo
Abstract
Southeastern Europe and, particularly, the Balkan Peninsula
are especially useful when studying the mechanisms responsible
for generating the current distribution of Paleolithic and
Neolithic genetic signals observed throughout Europe. In this
study, 404 individuals from Montenegro and 179 individuals from
Serbia were typed for 17 Y-STR loci and compared across 9 Y-STR
loci to geographically targeted previously published collections
to ascertain the phylogenetic relationships of populations
within the Balkan Peninsula and beyond.
We aim to provide information on whether groups in the region represent an amalgamation of Paleolithic and Neolithic genetic substrata, or whether acculturation has played a critical role in the spread of agriculture. We have found genetic markers of Middle Eastern, south Asian and European descent in the area, however, admixture analyses indicate that over 80% of the Balkan gene pool is of European descent. Altogether, our data support the view that the diffusion of agriculture into the Balkan region was mostly a cultural phenomenon although some genetic infiltration from Africa, the Levant, the Caucasus, and the Near East has occurred. Am J Phys Anthropol, 2010. © 2010 Wiley-Liss, Inc.
Streszczenie
(translated by Google)
Europa Południowo-Wschodnia, a zwłaszcza Półwyspu
Bałkańskiego są szczególnie przydatne przy badaniu mechanizmów
odpowiedzialnych za generowanie obecny podział paleolitu i
neolitu genetycznych sygnałów obserwowanych w całej Europie. W
badaniu tym 404 osób z Czarnogóry i 179 osób z Serbii. 17 loci Y-STR zostały
wpisane i porównywane przez 9 loci Y-STR do
ukierunkowanych geograficznie opublikowany wcześniej zbiory do
ustalenia filogenetycznych populacji w Półwyspie Bałkańskim i
poza nią.
Naszym celem jest dostarczenie informacji na temat tego, czy grup w regionie stanowią połączenie z paleolitu i neolitu podłoża genetycznego, czy akulturacji odegrał kluczową rolę w rozprzestrzenianiu się rolnictwa. Znaleźliśmy markerów genetycznych z Bliskiego Wschodu, Azji Południowej i pochodzenia europejskiego w tej dziedzinie, jednak analizy wskazują, że domieszka 80% na Bałkanach puli genowej jest pochodzenia europejskiego. Łącznie, nasze dane potwierdzają opinię, że rozpowszechnianie rolnictwa w regionie Bałkanów była głównie zjawiskiem kulturowym, choć niektóre genetyczne infiltracji z Afryki, Bliskiego Wschodu, Kaukazu i Bliskiego Wschodu miało miejsce. Am J antropologii Phys. 2010. © 2010 Wiley-Liss, Inc
Mój
komentarz. Głos powyższy to kolejny głos w dyskusji na temat
pochodzenia istotnego elementu życia rodzaju ludzkiego, jakim
jest rolnictwo. Pytanie brzmi: Kto je wprowadził w Europie?
Autochtoni, którzy z myśliwych-zbieraczy, przyjąwszy obce wzorce
kulturowe, stali się rolnikami, czy obcy przybysze, np. z
Bliskiego Wschodu. Ostatnio głosy się przechylają jednak ku
drugiemu stanowisku, zwłaszcza odkąd genetycy zaczęli sie
zgadzać na to, że zachodnioeuroopejska populacja R1b1b2 nie jest
autochtonna, lecz w okresie polodowcowym przywędrowała z Azji
Zachodniej, ze Wschodu, zapewne przez Anatolię w Azji Mniejszej,
podobnie zresztą jak i populacja R1a1a, której przybycie miało
sie dokonać nieco wcześniej prawie tym samym szlakiem, lub jak
twierdzili starsi genetycy - przez stepy nadczarnomorskie.
[6]
W. Haak et
al. 2010,
Ancient DNA from European Early Neolithic Farmers Reveals
Their Near Eastern Affinities.
http://www.plosbiology.org/article/info:doi/10.1371/journal.pbio.1000536
Jest to praca przełomowa, oczekiwana od dawna. Zespół genetyków, któremu przewodzi Wolfgang Haak, przeprowadził udaną analizę genetyczną 20 archeologicznych DNA: 17 mtDNA i 3 Y-DNA, z grobów cmentarza w Derenburg Meersentieg II w Saksonii. Metodą radiowęglową (skalibrowaną) pochówki datowano na czas 5500-4900 lat p.n.e. Groby i ich wyposażenie należą do kultury ceramiki wstęgowej (Linear Beaker Culture / Linear Bandkeramik, LBK). Wyjątkowość polega na tym, że jest to pierwsza identyfikacja Y-DNA, znanego z nietrwałości, z tak dawnego czasu. Poprzednie badania, zresztą bardzo głośne, doprowadziły do zidentyfikowania Y-DNA R1a1a trzech szkieletów z Eulau z roku około 2600 p.n.e.
Również ostateczny wynik jest zaskakujący. Zamiast bowiem spodziewanego R1a1a (jak w Eulau) w dwóch szkieletach zidentyfikowano haplogrupę F* (paragrupę) z grobu datowanego radiowęglowo z kalibracją na rok 5247 p.n.e. Haplogrupa F jest ojcowską dla haplogrup G, H oraz zespolonej IJK (przy czym nadto od K pochodzą haplogrupy: L, M, N, O, P, Q, R, S i T, które wyróżniają prawie 90% pozaafrykańskiej ludzkości). Haplogrupa F wstała około 48000 lat temu. Spotykana jest głównie w południowych Indiach na na Sri Lanka (Cejlon). Hg. F jest bardzo rzadką w Europie.
Natomiast w trzecim szkielecie, datowanym na rok około 5000 p.n.e., zidentyfikowano haplogrupę G2a3, która dominuje w rejonie Kaukazu, zwłaszcza w Armenii i Gruzji, oraz mniej w Turcji. W Europie bywa w Bawarii, Austrii (Tyrol) i Chorwacji (Osijek nad Drawą) 8-14%, w Zachodnich Włoszech i Sardynii 11-21%, w Turcji i na Kaukazie ogółem 11%.
Zidentyfikowano też dość liczne haplogrupy żeńskie, z których jedne ze względu na podwyższone lub wysokie częstotliwości są unikalne dla tej kultury na tle populacji przedneolitycznych łowiecko-zbieraczych i współczesnych, np. T, N1a, K i W (świadcząc o swoim pochodzeniu z LBK i o późniejszych trudnościach w przeżyciu), inna, haplogrupa H występowała wówczas w ponad dwukrotnie niższym niż dziś procencie w Europie (świadcząc o kolejnych migracjach późno neolitycznych), a wreszcie jeszcze inne znajdują zwykłą kontynuację w dzisiejszych populacjach (świadcząc o nieistnieniu pełnej wymiany populacji w czasach późniejszych).
Szczegółowe
porównania DNA z innymi kilkudziesięcioma populacjami Europy i
Azji Zachodniej wykazały największe podobieństwo do DNA krajów
Bliskiego Wschodu: Turcja, Kaukaz, Iran i bark tego podobieństwa
do DNA rejonu Wschodniej i Północnej Europy, tam, gdzie też nie
dotarła kultura LBK.
Ilustruje to mapa neolitu:
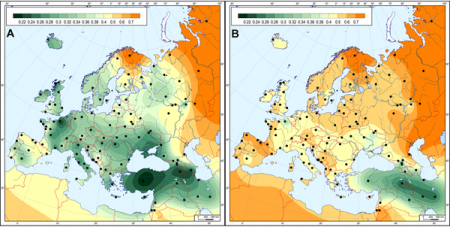
Na mapie A porównano dzisiejsze populacje całego
terytorium europejskiego neolitu LBK (punkty czarne oznaczają
miejsca porównywania dzisiejszej ludności). Kolor
ciemnozielony wskazuje na największe podobieństwo do populacji
bliskiego Wschodu.
Na mapie B porównano tylko populację LBK Denerburg.
Kolor ciemnozielony wskazał podobieństwo do bliżej określonego
regionu Wschodniej Turcji i Iranu.
Poniżej - mapa ekspansji neolitu środkowoeuropejskiego LBK i południowej kultury impresso (Cardium Pottery). Widoczne są dwie rzeczne trasy ekspansji: Dunaj i Dniestr.
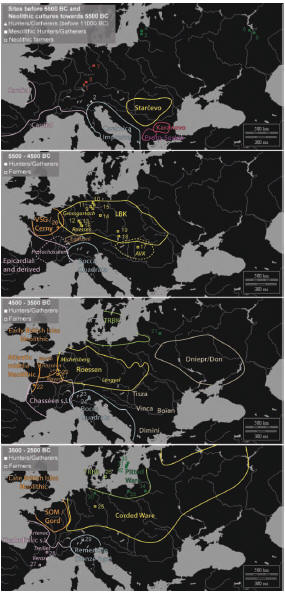
W komentarzach naukowych na antropologicznym blogu Dienekesa i DNA Forums podkreśla się zaskakujący brak haplogrupy R1a1a. Ale przecież wiadomo, że Derenburg Meersentieg II to zaledwie zaledwie jeden spośród kilkudziesięciu i więcej punktów osadniczych kultury ceramiki wstęgowej. Spodziewamy się, że badania kolejnych szkieletów z innych punktów osadniczych LBK ujawnią inne haplogrupy Y-DNA, zwłaszcza R1a1a.
[7] Projekt drzewa filogenetycznego haplogrupy R1a w FtDNA (Thomas Krahn)
z dnia 22.01.2011, po
odkryciu nowych SNP: L448, L449, L450, L451
http://ytree.ftdna.com/index.php?name=Draft
=R1a L62, L63, L120, M420, M449, M511, M513
= = R1a1 (może hg: R-M198) SRY10831.2, M448, L122, M459, M516
= = = R1a1a, M17, M198, M512, M514, M515, L168
= = = = R1a1a1 (może hg: R-M198) M417, PAGES00007, L449
= = = = = R1a1a1c M64.2, M87, M204
= = = = = = może hg: R-M458 L291
= = = = = może hg: R-M417* L12
= = = = = może hg: R-M417* L365
= = = = = może hg: R-M417* L366
= = = = = może hg: R1a1* PAGES00068
= = = = = może hg: R-M198 L448 (w gałęzi szkocko-skandynawskiej młodszej)
= = = = = = może hg: R-M417* L450 (w gałęzi północnokarpackiej)
= = = = = = może hg: R-M417 L451 (w gałęzi... ?)
Jak widzimy, pozycje M417 i M198 względem siebie są nadal
nieustalone. Nieustalone też pozycje nowych mutacji.
Konieczne są dalsze, liczne testy typu "Advanced".
Wykreślono problematyczny snip L170.
[8]
Koncepcje pochodzenia i migracji pozaafrykańskiej ludzkości,
w tym R1a i R1b, od haplogrupy F.
|
|
|
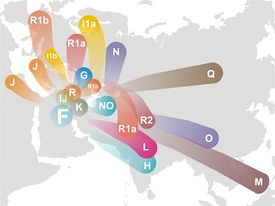
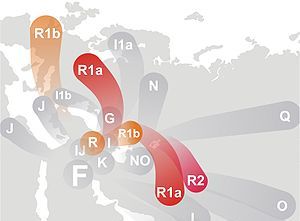
Obrzeża Morza Kaspijskiego jako ojczyzna haplogrup R, R1a i R1b.
|
DZIĘKUJĘ! |
|
GOŚCIEM |
Do strony Pietrzakowie. Genealogia.
Pietrzak
Do strony Kurczabowie. Genealogia.
Kurczab. Bołoz
Do strony Janikowie. Genealogia. Janik
Do strony Szlagowie. Genealogia.
Szlaga/Ślaga
Powrót do Strony
głównej witryny: Parafia TROPIE
Str.pryw.-genealog.
PIETRZAKOWIE.Pietrzak,
JANIKOWIE.Janik,
FIUTOWIE,
SZLAGOWIE Nadto:
Gierałt.GIERAŁTOWIE